|
|||
5. SISTEMSKA KOGNITIVNA NEVROZNANOST
FILOGENETSKI RAZVOJ BIOLOŠKIH STRUKTUR IN PREHOD K VIRTUALNIM STRUKTURAM
Že pri nižjih živalih (npr. hidroidnih polipih) se sprejem signalov in organizacija gibanja ustvarja z difuznim oziroma mrežastnim živčnim sistemom. Ni nobenega živčnega centra, temveč se "živčno težišče" seli iz enega v drugi del sistema. Z napredovanjem evolucije se je živčni sistem diferenciral. V prednjem delu možganov (v smeri premikanja) se se razvili specializirani prejemni (receptorni) sistemi nevronov, prednji gangliji pa so obdelovali dobljene informacije. Kasneje so se nevroni specializirali za izolirano zaznavanje vidnih, slušnih, kemijskih, tipnih, vonjalnih idr. dražljajev, za njimi pa se je razvila kompleksna funkcionalna struktura z asociativnimi in medprostornimi nevroni, ki sintezirajo informacije čutil. Tako je že pri vrtinčarjih in nadalje žuželkah idr. Pri vretenčarjih spremljamo razvoj centralnega živčnega sistema oziroma možganov: pri pticah vodijo medmožgani, ki že "sinergirajo"; pri sesalcih izstopi možganska skorja, ki že omogoča individualno-spremenljivo vedenje v dinamičnem okolju in s tem nadgradi prirojene vzorce. Kaže, da so se v evoluciji simetrične asociativne in plastovite nevronske mreže izmenično razvijale druga iz druge.
Pri človeku se (glede na psa) nekajkrat povečajo terciarni (to so asociativni) predeli velikih možganov – spodnjetemenski in čelni predeli. Vplivom naravnega okolja se dodajo še družbeno-zgodovinske okoliščine. Intenzivno se razvije sposobnost simboliziranja in kodiranja informacij, ki zaživi v jeziku kot dinamičnem psihofizičnem "organu-procesu" (Chomsky idr., 1990).
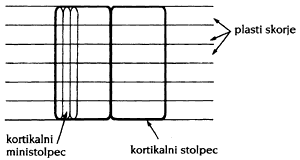
Kodiranje in sinteza informacij se torej zelo uveljavita. Nevronske mreže postajajo v višjih živčnih centrih vse bolj UNIFORMNE (Oakley, 1985; Ebdon, 1993) in gosto prepletene, torej asociativne. To bi v idealnem modelu pomenilo, da je vsak nevron enakovreden ter z vsakim drugim dvosmerno povezan (signali lahko potujejo v obe smeri). V realni mreži sicer niso vsi nevroni povezani z vsemi, vendar ni napak, da mrežo v približku modeliramo kar takole, ker so ODLOČILNI predvsem ATRAKTORJI! To dejstvo nas srečno rešuje iz zagat, ki jih prinašajo še neraziskane nevrofiziološke strukture. Zato je splošnost naše sistemske razprave obvarovana, ne glede na fiziološke detajle. Tudi matematično so dokazali, da razredčena mreža deluje precej podobno polnopovezani (Palmer idr., 1991 (str. 45–48), Peretto, 1992). V drugi interpretaciji pa so za osnovne dvosmerno interagirajoče elemente v simetrični mreži izbrani celi KORTIKALNI STOLPCI.
Za subkortikalne predele možganov so značilnejše večplastne nevronske mreže, v katerih potujejo signali najprej le v ENO SMER: od "vhoda" (senzornih sistemov) proti "notranjosti", kjer se informacije primerjajo in sintezirajo. Višje asociativne mreže so bolj simetrične oziroma uniformne in se specializirajo šele na nivoju vzorcev, ki jih oblikujejo. Tako so bolj prožne in prilagodljive. Nižje PLASTOVITE nevronske mreže (njihovi modeli so PERCEPTRONI: slika 16) pa so specializirane že na nivoju nevronov in samo sprejemajo dražljaje, jih razporejajo v razrede ter le delno obdelane pošiljajo dalje. Razliko uvidimo, če primerjamo sliko 7 in sliko 16.
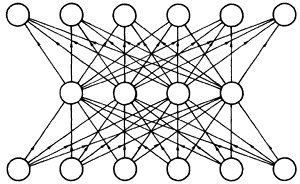
Slika 16. Perceptron – model večplastne mreže, kjer signali napredujejo
le v eno smer; brez
povezav znotraj istega nivoja
Srednja plast na shemi perceptronskega modela vsebuje kardinalne celice, ki ustrezajo nastajajočim razredom. V primarnih plasteh možganske skorje so odkrili (Lurija, 1983) na primer kardinalne nevrone, ki so specializirani za detekcijo pravokotnih ali samo fino zaobljenih linij, samo za ostre kote, samo za okrogle obrise, samo za gibanje točke od središča k periferiji ali samo od periferije k središču, za visoke ali za nizke tone, za specifične kinestetične, zvočne ali svetlobne dražljaje (npr. le pojav ali prenehanje svetlobnega snopa) idr. V sekundarnih plasteh skorje pa dobimo specifične odzive na še bolj zapletene lastnosti: na ritmične zvočne strukture, specifičen nagib, debelino ali konfiguracijo linije, na specifične vrste signalov ali na določeno število vidnih oziroma slušnih dražljajev itd.
Kot je vidno na sliki 16, nevroni v perceptronski mreži niso povezani med seboj v isti plasti, temveč njihove vezi vodijo le do celic višje plasti. Zato je perceptron manj prilagodljiv in bolj determiniran kot asociativna mreža, kjer so nevroni enakovredno povezani med seboj. Bolj determiniran pomeni, da je njegova funkcionalnost vgrajena v strukturo biološke mreže same. V perceptivnem aparatu se izvaja tudi LATERALNA INHIBICIJA (zatiranje) med sosednjimi nevroni, ki je dobro modelirana s KOHONENovimi mrežami (Kohonen, 1982, 1987). V teh mrežah se na specifičen dražljaj najbolj odzove specifičen nevron, potem ko je ob zatiranju sosedov zmagal v tekmovanju med nevroni (princip "zmagovalec dobi vse").
Pri asociativnih mrežah pa vsi razredi in opredeljene kategorije ne nastajajo na nivoju bioloških nevronov, temveč šele na ravni vzorcev kot konfiguracij nevronov. Asociativne sintetične strukture so torej virtualne oziroma "nadmaterialne"; so gestalti bioloških nevronov. Virtualni vzorci pa so seveda mnogo bolj spremenljivi, prožni in sintetični. Šele oni omogočajo vzrast človeka v duševno bitje kot velik evolucijski korak, ki ga je omogočil prehod od enosmernih plastovitih nevronskih mrež (perceptronov) samih k nadgradnji le-teh z bolj uniformnimi asociativnimi nevronskimi mrežami v možganski skorji. V zadnjem stavku smo mislili, kot vselej, na modele, ki najbolj ustrezajo dejanskemu živčnemu omrežju.
Simetrične asociativne nevronske mreže same in edine seveda ne bi bile primerne, saj ne bi bile sposobne hitre določitve in klasifikacije dražljajev iz hitro spreminjajočega se okolja. V poštev pridejo šele kot sintetična nadgradnja dodelane perceptronske analize. Taka kombinacija daje smisel tudi dediščini filogeneze oziroma dejstvu, da se možgani ne le funkcionalno spreminjajo, temveč predvsem nadgrajujejo.
Na virtualnem nivoju se tudi asociativne mreže v možganski skorji vzorčno diferencirajo v hierarhijo soodvisnih virtualnih struktur (funkcionalno podobno, kot je hierarhizirana primarna plast fiziološko). Pri primarni plasti skorje, kjer poteka zaznava (modelirano s perceptroni), smo torej govorili o FIZIOLOŠKI HIERARHIZACIJI, pri višjih asociativnih plasteh pa se razvije (tudi) VIRTUALNA HIERARHIZACIJA. To dogajanje najoptimalneje modeliramo s Hakenovim sinergetskim modelom. Treba je poudariti, da so možganska omrežja nevronov vselej kombinacija različnih arhitektur, ki se prepletajo na različnih velikostnih in funkcionalnih stopnjah. Po nekaterih kriterijih je takšnih možganskih arhitektur okoli 400, te pa se še dodatno sestavljajo in povezujejo v komplekse. Oblikujejo se povratne zanke, tako biološko-strukturne (posebne povezave med nevronskimi podmrežami) kot tudi funkcionalno-informacijske (signali določenega pomena med nevroni) in celo virtualno-informacijske (odnosi določenega pomena med vzorci).
Tukaj je treba še enkrat poudariti veliko kvalitativno spremembo, ki jo pomeni prehod od stanj posameznih nevronov (ki ponazarjajo neko informacijo, merjeno v bitih) prek vzpostavitve odnosov (vezi in korelacij) med nevroni do konfiguracij množice nevronov (ki odtlej predstavljajo gestaltno informacijo, merjeno v številu vzorcev). To je prehod od predstavitve modela oziroma sklopa informacij s stanji (vsebinami) nevronov k predstavitvi s konfiguracijami stanj, katerih posameznosti se zlijejo v vzorec-mozaik. Ta vzorec pa pomeni tedaj novo stanje – skupinsko stanje nove kvalitete, ki se stabilizira kot optimizirano stanje maksimalne usklajenosti nevronov oziroma minimalne energije sistema nevronov.
MOŽGANSKA SKORJA
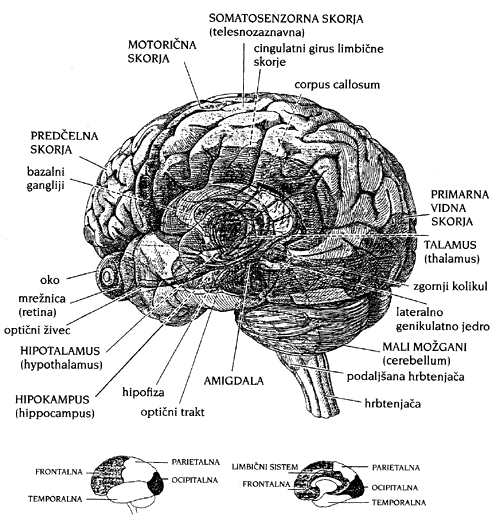
Najprej navedimo nižje dele živčnega sistema, ki sodelujejo pri organizaciji in regulaciji operacij možganske skorje. Gornji del možganskega debla in HIPOTALAMUS uravnavata homeostazo, ki zajema termoregulacijo, ravnotežje izmenjave materije, ki omogoča dihanje, hranjenje in obnovo, krmiljenje endokrinih žlez idr. Možgani so samoregulacijski sistem. V možganskem deblu je tudi RETIKULARNA FORMACIJA. To je zapletena mreža živcev, ki sprejema živčne signale iz raznih možganskih središč in jih razpošilja vsepovsod, vzdržuje budnost in filtrira informacije. HIPOKAMPUS razporeja spominske sledi (engrame) po možganski skorji (Vodovnik, 1991).
Možganska skorja in subkortikalne formacije sestojijo iz "sive substance" (nevronskih celic) in "bele substance". Slednjo sestavljajo aksoni, ki povezujejo posamezne predele skorje in jo spajajo s perifernimi in subkortikalnimi strukturami.
Prekinitev povezav med posameznimi področji možganske skorje ne povzroči nujno znatne spremembe v vedenju organizma. To dodatno nakazuje asociativno funkcionalnost in "medsebojno informacijsko vsebovanost" vsebin nevronov v paralelno-distribuiranem nevronskem sistemu ("vse v enem, eno v vsem"). Tako del mreže lahko asociativno obnovi delovanje celotne mreže ali ga nadomesti. To je razlog, zakaj celo težke poškodbe ali kirurška odstranitev asociativnih predelov možganske skorje ne uničijo nujno delovanja. Pač pa prekinitev povezav med možgansko skorjo in nižjeležečimi strukturami pogosto povzroči znatne motnje regulacijskih funkcij.
Nova možganska skorja (NEOKORTEKS) je navpično organizirana v šest plasti celic, od katerih se le spodnje povezujejo s periferijo. 4. sloj se veže s čutili, 5. sloj (ki vključuje gigantske piramidalne celice) pa z mišicami. Ločujejo se naslednja PROJEKCIJSKA SENZORIČNA območja: vidno (zatilno), slušno idr. (senčno) in območje splošne občutljivosti (temensko) (Lurija, 1983). Nanje se topološko korektno preslikajo polja receptorjev (nevronov v čutilih). Tem primarnim slojem sledita 2. in 3. sekundarna plast, ki ju sestavljajo celice s kratkimi aksoni in predelujejo informacije iz 4. in 5. plasti. Slednji sestavljata INTEGRATIVNE ali asociativne nivoje skorje, kamor spadata npr. Broadmanovi polji št. 39 in 40. Predstavljajo interpretativna področja, "kjer samozavedajoča duševnost vzpostavi stik z možgansko skorjo" (kot je dualistično rečeno v: Vodovnik, 1991).
Terciarni predel možganske skorje, ki ga sestavljajo mejne cone med kortikalnimi sekundarnimi območji, sintezira vse informacije iz podrejenih plasti. Terciarni predel je torej območje preseka (soprekrivanja) kortikalnih območij posameznih analizatorjev ter nosi najbolj zapletene integralne funkcije možganov. Sestavljajo ga izključno asociativne mreže, ki nimajo neposredne zveze s periferijo. Terciarni predeli se nahajajo na stikih zatilne, zadnjecentralne in senčne skorje.
Podobna je hierarhična organizacija motoričnih predelov, le da gredo operacije tudi v nasprotni smeri: signali potujejo iz opisanih višjih asociativnih plasti proti motorični periferiji. Pri tem pa se na osnovi posameznih impulzov v višjih motoričnih integracijskih plasteh sintezirajo sistemi gibov in sestavljenih kretenj.
Da v zgornjih predelih skorje prevladujejo asociativne nevronske mreže, dokazuje število povezav na število nevronov. Perceptronske mreže imajo manj povezav, saj le-te potekajo le od nižje k višji plasti, ne vežejo pa se nevroni iste plasti. Asociativne mreže pa imajo precej več povezav, saj se nevroni oziroma kortikalni stolpci vežejo med seboj enakopravno in v parih (v modelu vsak nevron ali kortikalni stolpec z vsakim drugim, v praksi pa večina z večino neposredno ali pa vsaj posredno). Da v višjih plasteh skorje, kjer vladajo asociativne mreže, domujejo višje duševne funkcije, dokazuje tudi ugotovitev, da ima otrok ob rojstvu že oblikovane subkortikalne formacije in najenostavnejše primarne predele, nima pa še dovolj razvitih sekundarnih in terciarnih plasti skorje. Nerazvitost se kaže v majhni širini oziroma funkcionalnosti teh plasti, majhni površini, ki jo zavzemajo, in v nerazvitosti celic, ki jih sestavljajo. Višji asociativni predeli se torej razvijajo šele z življenjem in učenjem. Šele pri opicah so terciarne strukture diferencirane od sekundarnih; pri drugih živalih to še ni opazno (Lurija, 1983). Tudi pri človeku je NEOKORTEKS PO ROJSTVU ŠE NEDIFERENCIRAN in funkcionalno nedoločen. Šele vpliv zunanjih dražljajev, ki jih skorji prenaša talamus, poganja rast in razvoj neokorteksa ter povzroča hkratno počasno virtualno diferenciacijo in specializacijo (Killackey, 1990).
Razmerje med maso glijalnega tkiva in maso nevronskih celic evolucijsko narašča. To kaže, da glija-celice niso le podporne celice nevronom, temveč imajo morebiti pomembno vlogo pri možganskih procesih (kot ojačevalniki?, koprocesorji?). Tudi razmerje dendriti : aksoni raste v korist dendritov, kar razkriva rastočo razvejanost dendritskega "vejevja".
SINTETIČNA DEJAVNOST SKORJE
Zadnja desetletja se stroga lokalizacija možganskih funkcij v določene predele vsaj za višje duševne funkcije opušča. Posebno to seveda velja za sestavljene funkcije. Lokalizirane so le osnovne funkcije, ki pa ne bodo predmet obravnave v tej knjigi, ker so za to na razpolago mnogi nevrofiziološki in medicinski učbeniki. Že običajno zaznavanje in predstavljanje predmeta je sinteza več osnovnejših zaznav. Nosilec sinteze so v filogenezi živčnega sistema vse bolj postajale specializirane in diferencirane funkcionalne strukture. Take strukture, če so fiziološke narave, so rezultat zelo dolgotrajne evolutivne samoorganizacije živčnega sistema. V višjih centrih pa gre za virtualne strukture, ki se razvijajo s kombiniranjem in superponiranjem vzorcev precej hitreje. Okolju dobro prilagojene strukture se zakodirajo v jakosti sinaptičnih vezi, da se ustrezna stanja nevronov in na njih temelječi vzorci ustalijo.
Prevladuje torej prepričanje, da gre pri možganskih funkcijah za globalne in kompleksne skupke vzorcev in procesov. V sestavljanju le-teh sodelujejo mnogi lokalni predeli in njihove sekundarne ter terciarne nadgradnje. Dražljaji (npr. vidni) se obdelujejo na večih nivojih. Groba diferenciacija in filtracija se opravi že na očesni mrežnici, s tem pa se omogoči npr. refleksni gib brez prejšnje zavestne ali višje možganske obdelave. V subkortikalnih vizualnih jedrih se razločijo npr. različne stopnje svetlosti. Sestavljene oblike vidnega opažanja se tvorijo v možganski skorji. Končno pa se sinestezije ustvarjajo v terciarni plasti skorje.
V otroštvu sta zaznavanje in pomnjenje osnovna procesa, seveda neodvisna od šele nastajajočih virtualnih struktur. Pri odraslem človeku pa postaneta kompleksna in tesno povezana z višjimi strukturami. Človek ne le opazuje in pomni, temveč tudi primerja, ocenjuje, se spominja in predvideva. V to pa so seveda vključene integrativne asociativne funkcije. Še "najenostavnejšo" oziroma najbolj hitro in elegantno možnost modeliranja dajejo spet simetrične asociativne mreže s prožno virtualno hierarhizacijo. Vzorci kot potencialne vrtače v konfiguracijskem prostoru nevronskega sistema so gnetljivi, dinamični ter zmožni "kompromisov" – zunanjih (ki jih sklenejo različni vzorci med seboj) in notranjih (vzpostavljenih med lastnimi konstitutivnimi vzorci nižjega reda). Te višje strukture, potem ko so se v duševnem dozorevanju človeka enkrat oblikovale, znatno pogojujejo tudi elementarne funkcije – s tem, ko jih povezujejo v razne kontekste in odnose. Odsekane in prvotno neodvisne poteze se zlijejo v izpiljene postopke – kompleksne gestalte.
Specializiranost struktur možganske skorje od primarnih k hierarhično višjim terciarnim plastem pada. Prav tako se s staranjem človeka težišče vse bolj prenaša k terciarnim strukturam, medtem ko primarne in sekundarne (denimo motorične) dejavnosti postajajo vse bolj avtomatične ali pa so pod nadzorom terciarnih struktur (npr. človek v pomembni situaciji pazi na "držo"). S tem se tudi nove zaznave tolmačijo v luči višjih asociativnih centrov ter v njih nastalih vzorčnih, simbolnih oziroma logičnih sistemov in posplošitev. Nastaja globalni kompromis in medsebojna kompenzacija med pričakovanji, predvidevanji, prepričanji in naravnanostjo (notranjim stanjem človeka) ter dejanskim zunanjim stanjem.
Čelni predeli možganske skorje poleg sinteze dražljajev ustvarjajo tudi programirane procese vzbujanja (v povezavi z govorno, miselno in zavestno-plansko dejavnostjo), nadzorujejo tok dejavnosti in beležijo njihovo izvršitev. Razen tega čelni predeli določajo usmerjenost k določenemu cilju gibanja. Ti terciarni prefrontalni predeli so nadgrajeni vsem delom skorje velikih možganskih polobel, in sicer ne samo tako kot terciarni predeli zadnjih delov skorje, ki pokrivajo npr. le dva sekundarna predela. Povezani pa so tudi z medmožgani in možganskim deblom. Tako lahko opravljajo najbolj univerzalno organizacijo splošnega vedenja človeka (Lurija, 1983).
VSEBINE RAZNIH PREDELOV MOŽGANSKE SKORJE
Rezultati poškodb in draženja posameznih mest v skorji
Pogosta metoda raziskav v preteklosti je bila sprva opazovanje vedenja ob slučajnih poškodbah možganov pri ljudeh, kasneje pa uničenje določenih središč pri živalih. Tako je poškodba določenega predela možganske skorje lahko povzročila motnjo njegove analitične in sintetične funkcije, ne da bi pri tem zmotili osnove zaznavnega procesa. Danes se nevropsihologi vse bolj posvečajo globalnim možganskim teorijam: npr. holografskim (Bohm, 1980) in holonomskim (Pribram, 1991), pri čemer skoraj vse temeljijo na zasnovi modelov nevronskih mrež, izhajajoč tudi iz omenjenih raziskav.
Predstavimo razne predele možganske skorje z njihovo vsebino, ki se je izrazila pri umetnem draženju teh predelov (Lurija, 1983). Draženje primarne projekcijske plasti vidne skorje s šibkim tokom so vzbujale videnja plamenčkov, pisanih peg, modrih diskov, svetlečih točk itd. Draženje sekundarnega vidnega dela skorje pa povzroča bolj figuralne privide: obraze, slike živali, rastlin, predmete itd., včasih pa tudi sestavljene scene (s strani prihajajoč znanec daje z roko nekakšen znak). Poškodbe sekundarnih plasti preprečujejo integracijo opažanja – sestavljanje posameznih vtisov v like in zaokrožene celote. Osnovni podvzorci se ne sintezirajo v vzorce – gestalte. Pacient normalno posebej zaznava posamezne dele in lastnosti, vendar raztrgano in nepovezano v enotno sliko. Na primer, gleda očala in ugiba: "Krog ... še en krog ... palica ... ne vem ... to mora biti kolo!" V drugem primeru lahko prepozna posamezne dele avtomobila, značilnosti in barve, ne ugotovi pa, da je to njegov avtomobil. Nejasnost povzročajo večpomenske besede, odvisne od konteksta: npr. v povedi "Gori na gori gori".
Poškodbe v terciarnih predelih skorje povzročajo na primer naslednje motnje: zamenjava pomena zaporedja besed (učiteljev prijatelj in prijateljev učitelj), nerazumevanje zapletenih stavkov, nezmožnost poimenovanja prepoznanega predmeta, dezorientacija v prostoru in na zemljevidu: zamenjava levo-desno, vzhod-zahod itd.
Ni pa prizadeto sekvenčno procesiranje (npr. zaporedje računskih operacij, plani in vrstni redi dejanj), temveč je ogrožena le zmožnost prostorske sinteze in hkratnih operacij.
Asociativni predeli so torej ustrezni za celovito vidno zaznavanje in sinteziranje. Logični in miselni procesi algoritmičnega tipa pa zahtevajo drugačne, ne nujno asociativne operacije, kjer je simetrija nevronske mreže zlomljena ter je dinamika sistema usmerjena in kaskadna. Takšno sekvenčno procesiranje lahko udejanjajo tudi virtualne strukture asociativnih mrež (s prehodi ali zveznim prelivanjem iz ene vzorčne konfiguracije, ki je le prehodno stabilna, v novo), vendar to ni nujno. S samoorganizacijo se lahko kodiranje ali zametki simboliziranja določenih vzorcev prenese na določene kardinalne celice ali (pravilneje rečeno) kardinalne domene v naslednji plasti hierarhije, ki pa potem interagirajo med seboj ne kot elementi paralelno-distribuiranih vzorcev, temveč kot nosilci lokalizirane informacije. To bi bil zametek logične aktivnosti z natančno opredeljenimi simboličnimi reprezentacijami in formaliziranimi procesi sklepanja, ki so značilni za raven umetne inteligence in govorne logike.
Leva in desna možganska polobla
Znane so razlike med desno in levo možgansko hemisfero. V primarnih slojih razlike še ni, le vlogi desne in leve strani se zamenjata. V sekundarnih plasteh pa se že izkaže, da je leva možganska polobla posvečena razvrščanju v kategorije, govoru, logičnemu, slovničnemu in analitičnemu razmišljanju, upoštevanju pravil itd.; desna polobla pa je namenjena celovitemu sintetičnemu mišljenju, bolj umetniškemu in intuitivnemu dojemanju. Pričakovali bi torej v desni hemisferi bolj asociativne strukture, v levi pa so le-te že močneje diferencirane in samoorganizirane v bolj trdne strukture, ki delujejo sekvenčno.
Pri asociativnih procesih gre za stapljanje vzorcev in s tem za pretvorbe navpično po hierarhiji vzorcev (npr. posploševanje, globalni vtis). Pri sekvenčnih procesih pa gre za vodoravne transformacije, torej pretežno na istem hierarhičnem nivoju (npr. naštevanje, "abecedovanje", uradni govor, zaporedno računanje). Vendar se je tudi pri sekvenčnih procesih treba zavedati asociativnih in kontekstualnih zvez ter vplivov, kar vodi do uporabe večplastnih asociativnih mrež. Takšne vsebuje na primer Hakenov sinergetski model. Tudi eksperimentalni podatki kažejo, da je funkcionalna organizacija v desni polobli precej manj diferencirana kot v levi. Procesiranje v levi polobli je torej bolj konkretne in formalizirane narave, v desni pa bolj splošno in svobodno.
Desna hemisfera je direktno povezana z zaznavnimi centri in omogoča bolj neposreden, naglo jasen, očiten, nenadzorovan in nezakodiran oziroma formalno nesimboliziran odnos do okolnega sveta. Desna polobla je bolj odgovorna tudi za samoopazovanje, samoidentiteto, samorefleksijo, za orientacijo v navadnem in v abstraktnih prostorih ter okoliščinah, za osebnostne lastnosti in nepojmovno zavest.
Zanimivo je, da človek določa številčnost neke množice pri majhnih količinah (npr. od 1 do 4 ali 5) nemudoma vizualno (vzorec zazna direktno), večje množice pa prešteva (z vzorci operira sekvenčno). Podobno napisane znane besede (npr. svoje ime) prepozna takoj kot en ustaljen vzorec. Manj pogoste besede pa bere po črkah in jih šele nato sestavlja v gestalt ter jih asociativno sproti popolnjuje. Od tod tudi zamenjave npr. "dežnik" in "deževnik" glede na kontekstualno pričakovanje.
Omenimo še pomembno dejstvo, da lahko ena možganska polobla nadomesti funkcijo druge, če je ta onesposobljena ali se ni razvila (Russel, 1987). V splošnem se lahko funkcije uničenih ali kirurško odstranjenih večjih predelov možganov sčasoma obnovijo, ker drugi prevzamejo njihovo vlogo. To je POMEMBEN DOKAZ, DA MOŽGANI DELUJEJO PRILAGODLJIVO, PARALELNO-DISTRIBUIRANO (torej NELOKALIZIRANO) in predvsem HOLISTIČNO (v smislu principa "vse v enem, eno v vsem" glede vsebovanja informacij in realizacije funkcij).
Obnovitev je lahko na fiziološki in še bolj neposredno na asociativno-virtualni ravni. Pri slednji je razlog v tem, da mreža lahko IZ DELA ASOCIATIVNO REKONSTRUIRA CELOTO, ČE SE POTENCIALNA VRTAČA ŠE NI ZABRISALA ob izgubi večjega dela elementov vzorčne konfiguracije (ki so jih predstavljali izgubljeni nevroni).
NAVEZAVA NA NEVROFIZIOLOGIJO
Zgradba nevronov, sinaps; nevrotransmiterji, nizi signalov
Izkaže se, da tudi (večkrat še neraziskane) NEVROFIZIOLOŠKE IN FUNKCIONALNE PODROBNOSTI NE SPREMENIJO OSNOVNIH PRINCIPOV DELOVANJA VZORČNIH VIRTUALNIH STRUKTUR v asociativnih predelih. Konfiguracije in potencialne vrtače nevronskega sistema se lahko oblikujejo na tak ali drugačen način. Edini pogoj je, da so informacijske preslikave in pretvorbe ENOLIČNE: specifičen vhod da vedno specifičen izhod – injektivno ali surjektivno (torej kategorizacijsko). Pri nevronskem sistemu je položaj podoben kot v sociologiji, kjer sociolog navadno sme zanemariti anatomijo posameznega človeka v svojih globalnih teorijah družbenih organizacij; mora pa pogosto upoštevati psihologijo posameznih ljudi. Pri nas temu ustreza stanje posameznih nevronov (aktivno/neaktivno) in sinaps (različne moči). Pri VELIKIH POPULACIJAH pa lahko zanemari tudi to in uporablja le tako imenovane teorije polja. Za modeliranje višjih duševnih procesov, v katerih sodeluje ogromno nevronov, njihovih skupkov, podmrež ter omrežij podmrež, lahko uporabljamo tudi samo metode fizike kompleksnih sistemov oziroma sinergetike ter njune matematične modele.
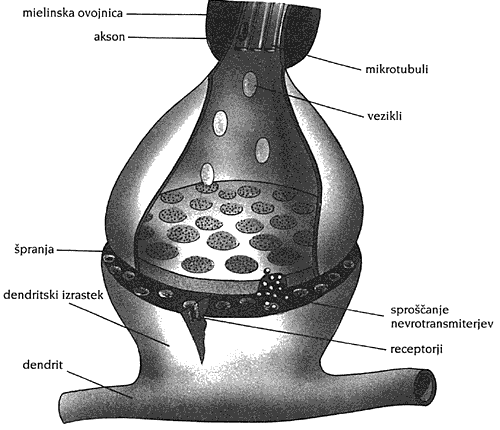
Vseeno si moramo ogledati osnove nevrofiziologije. Možgani vsebujejo približno 100 000 000 000 nevronov raznih vrst in okoli 100 000 000 000 000 sinaps. Na sliki 2 smo videli nevronove dele: telo živčne celice (soma), mnogo krajših (do nekaj mm) dendritov, en daljši (0,1 mm – 1 m) akson in sinaptične stike. Na sliki 17 je upodobljena sinapsa. Na eni strani stika so sinaptični mehurčki (vezikli), na drugi pa receptorji nevrotransmiterjev. Vmes potujejo NEVROTRANSMITERJI – kemične snovi, ki prenesejo signal od predsinaptičnega nevrona k posinaptičnemu nevronu. Kvanti (paketi) nevrotransmiterjev se izločijo iz sinaptičnih veziklov, ko električni signal po aksonu pride do njih. Prevladujejo sinaptični stiki akson-dendrit, obstajajo pa tudi stiki akson-soma, dendrit-dendrit, akson-akson.
Nevrotransmiterjev je okoli 30 vrst (Dobnikar, 1990). To so npr. acetilholin, norepinefrin in razni peptidi. Nekateri so vzbujevalni in prispevajo k vzburjenju nevrona. Tedaj le-ta začne oddajati nize sunkovitih signalov. Drugi nevrotransmiterji so zaviralni in preprečujejo vzburjenje nevrona. Vsak nevron sešteje prispevke drugih nevronov skozi zaviralne ter vzpodbujevalne sinapse. "Predznak" sinaps podaja narava nevrotransmiterjev. Če je vzpodbujenih signalov več kot zaviranih ter je še dodatno presežen določen prag, se nevron vzburi in oddaja vrste sunkov ("spike"-ov oziroma "akcijskih potencialov"), dokler je vzdražen.
Nevron deluje kot nelinearni pragovni element, kot ojačevalnik ter predvsem kot prostorski in časovni integrator (seštevalnik signalov iz določenega prostora v določenem časovnem intervalu). Deluje po načelu "vse ali nič": če zbere tolikšno količino signalov, da preseže prag, zapovrstno oddaja signale, sicer miruje. Nevroni delujejo čisto stohastično (naključno), analogno seštevajo signale in jih razpošiljajo asinhrono (ne vsi hkrati, temveč enkrat eden, enkrat drugi, kakor nanese). Edina diskretna stvar je prag, ki je presežen ali ne, in s tem tudi stanje nevrona, ki vzburjen ali nevzburjen.
Omenjali smo že Hebbovo učno pravilo, ki se v modelih redno uporablja. Opišimo sedaj natančno nevrofiziološki mehanizem, ki je izvor obrazca (2):